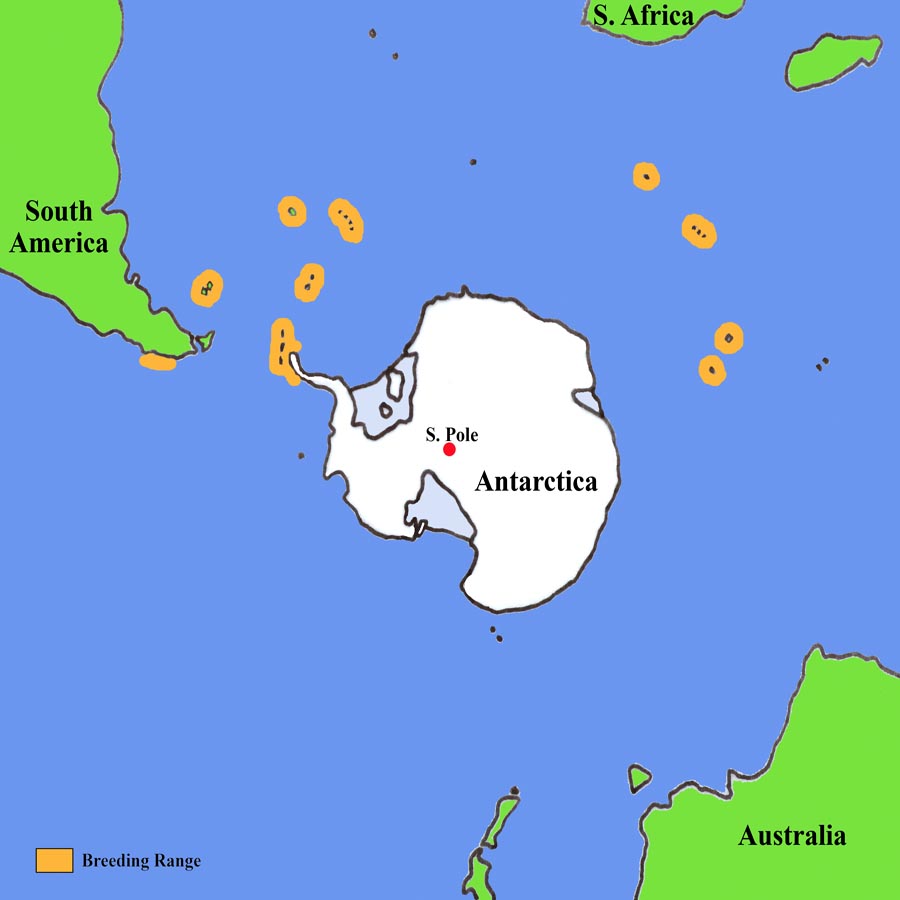
Nest and Partner Selection
Macaroni Penguins nest in large dense colonies often located on sloping ground above small cliffs. Compared to Rockhopper Penguins, with which they may establish mixed colonies in places, Macaronis nest on slightly more open ground, less often seeking the immediate shelter of rocks. Colonies are usually free of vegetation due to the wear on the vegetation imposed by the nesting penguins, although the penguins will also nest in surrounding areas of tussock grass. The colony is usually connected to the sea by one main walkway which may include extremely steep and rocky sections which the penguins are adept at mastering. The use of a single walkway facilitates monitoring of the transit of birds to and from colonies since it allows the use of automated gateways which detect passage of birds implanted with tiny passive transponders (C. Green et al., 2006. Polar Biol. 29(12), p.1993-1010). Penguins may spend a significant time transiting from the coast to their colony, for example 30 min each way at the Fairy Point colony, Bird Island, S. Georgia (Barlow and Croxall 2002. Ibis 144, p.248-258).
 |
 |
 |
|
Macaroni Penguin Colony, Elsehul, S. Georgia
|
Elsehul Macaroni Colony, Walkway
|
Elsehul Macaroni Colony. Inset: Penguins on walkway
|
The breeding season begins as birds return to their colonies after their extended winter foraging trips. Mass of penguins on arrival may be a critical determinant of breeding success (Crawford et al., 2007. Afr. J. Mar. Sci. 30(1), p.185-188; Raya Rey et al., 2007. Ibis 149, p.826-835). Male birds return first to establish nest sites, whereas females return about a week later. Nests are relatively simply and usually consist of little more than a slight hollow or a small pile of pebbles which may be lined with small amounts of plant material. The male is primarily responsible for collecting any nesting material, whilst the female tends to make adjustments during the egg-laying phase.
Fidelity to nest site and partners has been studied from 1986 to 1990 at Bird Island, South Georgia (Williams and Rodwell 1992. Condor 94, p.636-645). Nest site fidelity ranged from 69-87%, with mate fidelity between 74 and 78%. Separation was observed in 4-13% of cases, with non-return (probable death) of at least one of the mates accounting for the remainder (up to 19%). Successful breeders were less likely to change mate, yet subsequent breeding success was not higher in repeat breeders. It appears that males consider maintenance of nest site of primary importance. Males are reluctant to leave their nest sites to find a new partner and about 40% of males do not breed for at least 1 year following separation. However, females will find a new mate, usually within a few meters of the old nest site, in 94% of cases. Also, after separation in the case when both birds are alive, all males studies remained at the nest site, whilst the females established themselves at nearby nests. The high partner fidelity observed is probably accounted for by the high nest fidelity, which provides a focal point for re-pairing, together with the highly synchronized arrival of Macaroni Penguins at the breeding site, which allows little time for partner change.
Interestingly, about half of those birds whos partner did not return to moult already established a new partner during the moult period. Of these new partners, about half were actually mated with in the subsequent season. Hence, the moult appears to be an important period for establishing new pair bonds.
Courtship and Copulation
Courtship behaviour involves advertising such as bowing or the ecstatic display, in which eudyptid penguins point their heads skyward, extend their flippers and make a series of cries with wide open bills as they swing their heads from side to side. As courtship progresses, the pair may perform mutual displaying where both partners bow, quiver and perform ecstatic displays synchronously.
Copulation occurs at the nest site and may be instigated by either partner. The female may indicate willingness by squatting on the nest or the male by beating the female with one or both of his flippers. The male huddles up to the female, nibbles her neck gently and pats her with one of his flippers. The male then eventually mounts his partner who lies in a position with the head raised, and then edges backwards approaching the cloaca whilst using his flippers to pat the female's flanks. Both partners bring their cloaca towards each other so that contact can be made. The male briefly holds his position, stabilized by his flippers and usually also by the female who holds his neck with her bill. The male finally dismounts and adopts a subdued hunched posture. Both birds may proceed to preen or rest for some time following copulation.
Laying / Brood Reduction Mechanism
First eggs are usually laid by the end of November in S. Georgia. This time may fluctuate slightly between years or site. Two eggs are laid over a period of about 4 days, a smaller (A) egg, followed by the larger (B) egg. Egg composition studies revealed that the yolk of the (A) egg is proportionally larger (although smaller in absolute terms) than that of the (B) egg (30 v 24%), with the albumen (white) being accordingly smaller (57 v 63%), and the egg shells making up the rest (12 v 13%) (Gwynn 1993. Emu 93, p.290-291). It should be noted that it has been suggested by one author that the (A) egg is simply smaller since it develops during a period when yoke deposition is below maximum (Williams 1990. Oikos 59(3), p.349-354).
All crested penguin species display egg dimorphism and associated brood reduction mechanisms, favouring the second and larger (B) egg. There are however significant differences in the extent of dimorphism and the kind of brood reduction behaviour displayed. Macaroni, Royal and Erect-Crested Penguins show the most extreme form of brood reduction and appear to actively remove the first laid (A) egg from the nest, whilst the Rockhopper, Fiordland Crested and Snares Penguins do not actively remove the (A) egg and may in rare cases even raise 2 chicks. The Macaroni Penguins eggs are the second most dimorphic of those found in the Eudyptid Penguins, only surpassed by those of the Erect-Crested Penguin. On Bird Island, S. Georgia, the first (A) egg and second (B) egg having respective mean masses of 94 and 145 g, and a mean difference in size of 60% (although varying from 20% to 90% in different clutches) during the 1998 breeding season (Barlow 2001. Antarctic Sci. 13(3), p.286-287). An earlier study at Marion Island had found the mean size difference to be about 62% with hardly any variation, whilst the sizes of both eggs compared from one clutch to another fluctuated by up to 73%, as opposed to about 40% in the Barlow study (Williams 1990. Oikos 59(3), p.349-354).
Loss of the (A) egg occurred in 54% of nests prior to laying of the (B) egg during a study of Macaroni Penguins on Marion Island (Williams 1980. Auk 97, p.754-759). Similar figures were found in a later study which showed peak loss on the day before laying of the (B) egg and on the same day. All (A) eggs were lost by the end of the day on which the (B) egg was laid (Barlow 2001. Antarctic Sci. 13(3), p.286-287). Studies on the similar Royal Penguin show that eggs are removed from the nest by use of the bill, or are scraped out of the nest with the feet. This usually coincided with a period of increased parental nest-building behaviour preceding laying of the second egg (Cassady St Clair et al., 1995. Animal Behaviour 50, p.1177-1185). The reasons for this extreme form of brood reduction involving expulsion of the in principle viable (A) egg at such an early stage are not clear. Egg expulsion apparently unnecessarily limits the (A) egg insurance value to the 4 days preceding laying of the (B) egg. It is possible that the (A) egg primarily serves as an insurance against the failure to lay a second (B) egg. In studies where (A) eggs were artificially substituted for (B) eggs in some nests, and subsequently naturally incubated until hatching, it was found that (A) chicks had on average 55% of the mass of (B) chicks at hatching (Williams 1990. Oikos 59(3), p.349-354). By fledging, (A) and (B) chicks had the same sizes, and mass was similar or only marginally lower in (A) chicks. Hence, (A) eggs evidently have a similar potential to (B) eggs.
Experiments have shown that if a second egg (transferred from another nest) is added to the nest shortly after laying of the (A) egg, loss of the (A) egg is not accelerated. In fact, both the (A) egg and transferred egg were removed from the nest as usual within 24 hours of laying the (B) egg (Barlow 2001. Antarctic Sci. 13(3), p.286-287). In the same study it was shown that a transferred egg added to the nest after laying of the (B) egg was retained for an average of 6 days and was always incubated in the anterior position, which is preferentially taken by the smaller (A) egg in other Eudyptid Penguin species that incubate 2 eggs. Hence, Macaroni Penguins will not rapidly eject a second egg from the nest after the (B) egg has been laid. The fact that two eggs can clearly be incubated by Macaroni Penguins makes it even more mystifying why the (A) egg is lost so early.
Incubation
Incubation duties are divided into two main shifts, which are preceded by a phase where both parents perform a number of short alternating foraging trips. When a partner returns to the nest, the partners usually greet each other. This can involve loud trumpeting calls which may be performed with the head pointing upwards and the flippers flapping up and down, and/or may include some head-swinging as in the ecstatic display. If nest relief occurs, the partners adopt a relatively hunched posture and the relieving bird shuffles slowly onto the nest, shifts from foot to foot a couple of times and then gently settles onto the nest. When the main incubation shifts start, the male leaves to forage and regain weight whilst the female remains on the nest. This period lasts for about 10-20 days. The remainder of the incubation duties are then largely left to the male, whilst the female forages (Williams and Croxall 1991. J. Zool. (London) 223(2), p. 189-202). During the male incubation shift, females (if foraging locally) may sporadically return to the nest and perform brief incubation shifts (mean 10 h) whilst the male stands nearby (Haftorn 1986. Polar Res. 4 n.s. p.33-45). Interestingly, incubating males were observed to spend about 43% of their incubation duties in an upright position (including 3% of time preening, displaying, shifting the egg or turning on nest) and 57% in the prone (lying) position. The proportion of time spent upright is exceptionally high for an incubating penguin. It appears that it is possible to effectively incubate a single egg clutch effectively without adopting a prone posture, since egg temperature measurements revealed temperatures of over 37'C in the incubating birds.
Brood / Guard Phase
As in other Eudyptid penguins, after hatching of the egg, the male Macaroni Penguin remains on the nest and has sole responsibility for guarding the chick, whilst the female provisions the chick by making short 1-2 day foraging trips. Chicks hatch with a thin coat of down (protoptile plumage) which allows efficient heat transfer from the brooding parent. A thicker down (mesoptyl plumage) is developed within 2-3 weeks. Young chicks are brooded under the parent, but rapidly become too large to fit under it. Large chicks lie partially covered (usually with head and shoulders covered) by the parent or lean closely against it. The mesoptyl down is crucial for thermoemancipation of the chicks, and is a prerequisite for the transition from the guard to the creche phase which usually occurs after about 20-23 days at Bird Island. Once the chick is in the creche phase, both parents provision the chick. Creching is thought to provide some protection against predation. When parents return, the chicks need to be recognized so that they can be fed.
Creche Phase / Acoustic Parent-Chick Recognition
During the creche phase, the chick no longer remains at the nest and is usually found about 3-5 m away from it in a small group of chicks called a creche. Adult-chick recognition now relies on acoustic signals. When arriving at the nesting site, adults make several about 1.75 sec long calls, made up of a long 1st part (syllable), followed by a second part of similar length consisting of a series of 5-30 (av. 12.84) shorter syllables (Searby et al., 2004. Animal Behav. 67, p.615-625) of which the first are repetitive short syllables, with the last 2 syllables being slightly longer. The pattern (or tempo) of the syllables, together with the harmonic content, make the signal distinctive, i.e. Macaroni Penguins have a double vocal signature. The call is fundamentally similar to, but significantly longer than that of the Rockhopper Penguin (0.45 sec), and the individual syllables are more distinctive in the Macaroni penguin (Searby and Jouventin 2005. J. Avian Biol. 36, p.449-460).
Both of these Eudyptid penguin calls are less complex than the double vocal signature of the Aptenodytes penguins (i.e. Emperor, King), but more complex than the solely harmonic system in Pygoscelis penguins (e.g. Adelie, Gentoo). The higher complexity of the call in non-nesting penguin species is accounted for by the fact that the acoustic signal does not only have to distinguish between birds at a particular location (i.e. nest and immediate surroundings) within a colony, but has to be able to discriminate essentially all birds in a colony without a supplementing positional signal. Playback experiments of Macaroni parent calls were able to stimulate chicks to return to the nest in 39% of cases, whereas a parental call was responded to in 99% of cases. This confirms that a visual stimulus of a parent at the nest is also important. Further, by modifying calls, it could be shown that the second part of the call carries most recognition information, especially encoded by the duration of the repetitive syllables. On hearing a parental call, chicks call and approach the parent, after which feeding usually commences within less than a minute. Parents recognize their own chicks acoustically and peck other chicks that approach them in the hope of being fed.
Provisioning of Chicks
Based on studies using implanted data loggers which were able to record heart rate and dive depth for Penguins over a full year, it was calculated that only about 15% of prey consumed during the chick-rearing phase was actually provided to chicks by both male and female birds (Green et al., 2007. Mar. Ecol. Prog. Ser. 346, p.299-312). This could be broken down into brood phase (females provide 22.3% of food to chick) and creche phase (female 16.7% and males 10.2%). On an average foraging trip, most food caught near the beginning of the trip is digested by the adult. Food caught towards the end of the trip is brought ashore in a relatively undigested state (it is thought that penguins can downregulate their digestive activity) and fed to the chick as necessary, with the remainder then being digested by the parent on land. In the study, returning parents were thought to have only about 36% (female) and 42% (male) of their daily catch in their stomachs, whilst the rest had already been digested. Of this, the relative proportions digested by chick or parent vary depending on the stage in the chicks development. As the chick grows, its demands initially increase rapidly until levelling off and even sometimes dropping slightly shortly before fledging. Hence, the female needs to provide the chick with a progressively higher proportion of food (eventually nearly all) until the male joins her in foraging at the beginning of the creche phase. Females tend to combine numerous short trips with a number of longer ones during the creche phase and arrival and departure times become more erratic than during the brood phase, where they are largely concentrated at dawn and dusk (C. Green et al., 2006. Polar Biol. 29(12), p.1003-1010). This pattern had been observed previously and it has been suggested that the longer trips serve to allocate resources to the parent (Barlow and Croxall, 2002. Ibis 144, p.248-258).
Studies on provisioning behaviour at Bird Island, S. Georgia in the 1998-2000 breeding seasons detailed the relative contributions of male and female birds and also chick development (Barlow and Croxall 2002. Ibis 144, p.248-258). On average, females provided 44 meals to the chick (17 during the 23 day guard period, 27 during the 37 day creche period) and males 19 (all during creche). Hence, even during the creche period, females accounted for 60% of meals, and in some individual cases males hardly fed the chick at all. Birds tended to leave the colony at dawn and return over a broader time-period leading up to dusk, with trips lasting 16 h on average. Birds spend an average of about 14 h on land between trips, males tending to stay about 2 hours longer. A minor peak in trip length records was also found at 42 h, corresponding to largely female birds spending a night out at sea and returning the next evening. Over the 3 study seasons, males provided approx. 0.5 meals/day and females 0.67. The male starts the creche period at a lower weight than the female and thus needs to focus more on its own condition. Male foraging rate is correlated with the chicks rate of growth and fledging mass. Females appear to perform a mixture of short and long trips, and it has been suggested that this may serve to feed the chick and maintain own condition, respectively. Average meals were about 300 g with significant interseasonal fluctuations presumably resulting from fluctuations in prey availability. For example, meals during the creche phase averaged 191 g in 1999 (when provisioning was also less frequent), but 391 g, in 2000. Irrespective of this massive difference in food provided to the chicks, there were only small differences in growth rates between years. Chick growth is nearly linear for the first 40-50 days, after which a mean weight in the region of 3 kg is reached, and then becomes more erratic shortly before fledging. Creching occurred at about 22 days at a mass of between 1-1.7 kg (mean approx. 1450 g). Mean fledging mass was 3.3 kg in 2000 and only 2806 kg in 1999, when food was more scarce. Fledging usually takes place after about 60 days on Bird Island. If the male does not contribute meals during the creche phase, chicks may still survive to fledging but are likely to only fledge at a mass of about 2400 g. The level of male contribution thus largely determines the mass at fledging, which is probably critical for survival during the first year (Barlow and Croxall 2002. Ibis 144, p.248-258). In the study, breeding success varied from 45.3% in 1998 to 58.9% in 1999 (even though chicks received least food in this year). This is largely due to the failure of 42.2% of incubated eggs to hatch in 1998, possibly due to poor female body condition upon arrival at the colony in this season. On average over the 3 years, 28% of nests failed due to hatching failure, 22% failed during the guard phase and only 4.5% during the creche period.
Chick food consumption totals approx. 17 kg between hatching and fledging. When enough food is available, the chick consumes about 110 g/day (i.e. equivalent to its own mass) shortly after hatching. This gradually increases and ideally peaks at about 400 g/day after about 30 days before then gradually declining again to about 180 g/day just before fledging. The mean daily time spent submerged for parents follows a similar pattern (Green et al., 2007. Mar. Ecol. Prog. Ser. 346, p.299-312). Males were found to be more efficient foragers, extracting 2.84 g (as opposed to 2.28 g for females) from their environment for every minute spent submerged foraging. Only about 15% of prey caught during the chick rearing period is actually fed to the chick, meaning that total parental consumption is about 110 kg during this period. Daily prey consumption could reach as high as 48% of body mass in males and 41% in females.
The data from Bird Island is certainly not representative for all years or study sites, and indeed appears to represent a situation with relatively good foraging conditions. In a study at Marion Island, chicks were observed fledging at an average of 2.3 kg at an age of 70 days, due to lower food availability (Brown 1987. Polar Biol. 8, p.95-102). Also, a study on chick-rearing Macaroni Penguins at Heard Island, Capsize Beach Colony, in 2003-2004 revealed average meal sizes of only 71 g and on average longer trips and thus less frequent meals.
Fledging
Macaroni Penguins fledge after approx. 60 days at Bird Island and have usually attained a mass of about 3 kg by this time. Earlier studies at Marion Island revealed fledging after 70 days at a lower mean weight of 2.3 kg (Brown 1987. Polar Biol. 8, p.95-102). The water-proof juvenile plumage develops shortly before fledging and lacks the distinctive crest of the adult birds. Juvenile penguins have a small crest at each side of the head, brown (as opposed to red) eyes and greyish-brown (as opposed to orange) bills.
Fledglings spend their first year at sea after which they return to land to moult into their adult plumage. Macaroni Penguins usually only start to breed at an age of about 5 years.
Post-Breeding Moult
After the chicks have fledged, adult birds go on extended foraging trips for at least 2 weeks to gain weight before the annual moult. During this period, penguins may accumulate significant reserves of lipids and proteins and thereby increase in mass by 50%, whilst at the same time the new feathers start to grow under the skin. Birds may arrive on land with from about 20-40% of the length of the new feathers already grown. The moult is necessary, since feathers suffer wear and need to be occasionally replaced to ensure that the penguins maintain a water-proof and insulating plumage. During the moult, penguins are relatively sedentary and remain ashore fasting. The moult has long been considered highly energetically demanding, but a detailed study on the energetics of moulting female Macaroni Penguins on S. Georgia have shown that energy expenditure may not be higher than during the breeding season (Green et al., 2004. J. Avian Biol. 35, p.153-161). In this study, the average length of the moult was 22.3 days, although based on other studies, the moult may take anything from 13-34 days. As the moult progressed, body mass loss was more or less linear in absolute terms, whilst the mean heart rate (measured using an implanted logger) increased until peaking after the moult was 60-70% complete, after which it slightly dropped again. A similar pattern was observed for the abdominal temperature. At this stage, metabolic activity is elevated to compensate for surface heat loss. This is due to the almost complete loss of the old plumage, the high level of vascularization of the skin, and the fact that only about 50% of the length of the new feathers have emerged.
After completion of the moult, the penguins go on their long winter foraging trips during which they do not return to land until the start of the next breeding season.
 |
 |
 |
|
Macaroni Penguin Incubating in Prone Position
|
Macaroni Penguin with Chick
|
Macaroni Penguin Inspecting Chick
|
 |
 |
 |
|
Macaroni Penguin with Chick
|
Macaroni Penguin with Chick
|
Macaroni Penguin with Chick
|
 |
 |
 |
|
Macaroni Penguin Chick
|
Macaroni Penguin with Chick
|
Macaroni Penguin Chick
|
|