Northern Rockhopper Penguin |
|
Specific Name: Eudyptes moseleyi | ||
|
|
|||
| Adult Height: 45-58cm | |||
| Adult Weight: 2.5-3.5kg | |||
| Adult Flipper Length: 17.5-19cm | |||
| Estimated Population: 350000 breeding pairs |
Distribution: |
||||||||
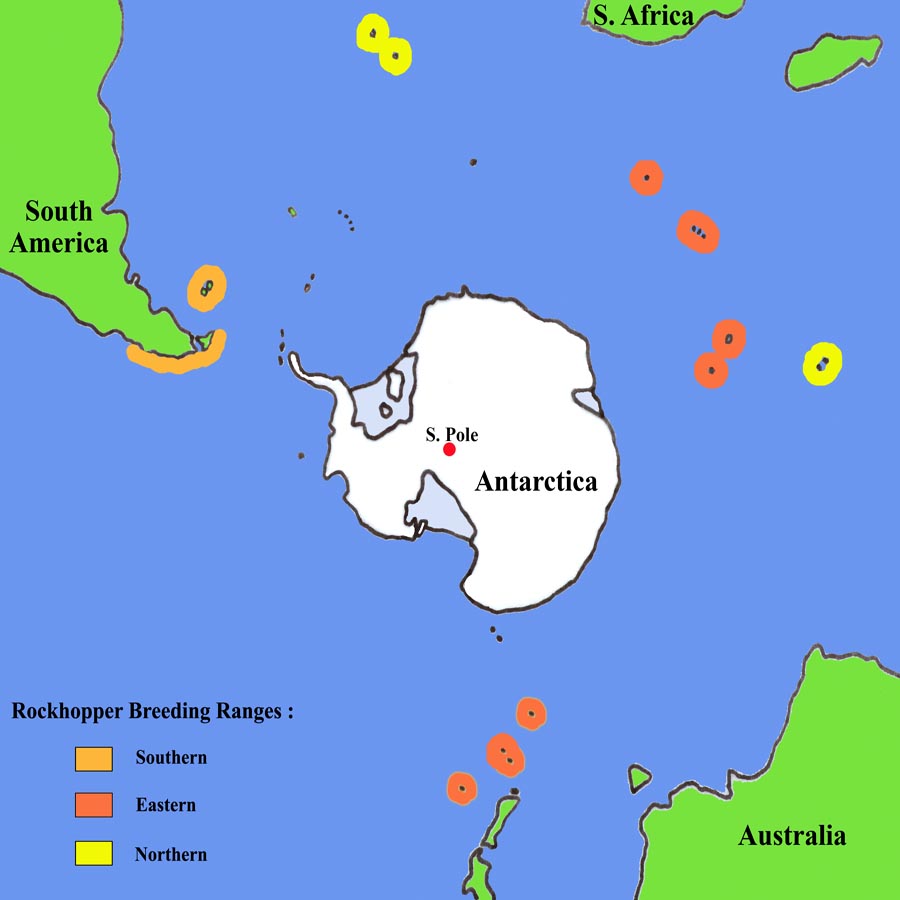
Northern Rockhopper penguin breeding sites are found on Tristan da Cunha, Gough and the Amsterdam & St Paul islands as indicated on the map. Due to their notably longer-crests compared to the other Rockhopper species, they are also known as Long-Crested Rockhoppers. The northern species is the only Rockhopper to breed in sub-tropical waters.The reader is also referred to the section on Rockhopper Penguins in general, since much of the information obtained in the more numerous studies on the other Rockhopper species also applies to the Northern Rockhopper population.
Separation into Northern and Southern species has been long suggested based on differing crest and song characteristics, both of which are relevant to breeding (Jouventin 1982. Visual and Vocal Signs in Penguins, their Evolution and Adaptive Characters. P. Parey Ed. Berlin). Genetic studies by the same author based on differences in the mitochondrial control region and ND2 genes between Rockhoppers from N and E (referred to as S in paper as S/E populations considered to be same), confirmed that at least a N and a S population could be distinguished (Jouventin et al., 2006. Mol. Ecol. 15, p.3413-3423). The species were estimated to have split as little as 140000 years ago, but more certainly less than 1 Million years ago (inaccuracy as rate of DNA mutation upon which calculation is based can only be estimated).The further separation into 3 species was based on analysis of differences in sequences in the small ribosomal subunit (12S), cytochrome B, and cytochrome oxidase subunit 1 genes between populations from N, S and E areas (Banks et al., 2006. Polar Biol. 30, p.61-67). These differences were considered sufficient to merit separation into three species. The N species is genetically less related to the S and E populations than the latter pair to each other. In fact, the N and S genetic divergence was higher than that between Galapagos and Humboldt penguins (two recognized species) in two out of the three genes analysed. The Northern species is also easily distinguishable from the other two based on morphological features, whereas S and E species are visually indistinguishable. For example, the Northern Rockhopper has a far longer crest, a larger area of black plumage on the tip of the underside of the flipper and a thick black leading edge of the flipper (the other species have a thin grey leading edge on the underside of the flipper). Northern Rockhoppers are also marginally longer bills and flippers (Tremblay and Cherel 2003. Mar. Ecol. Prog. Ser. 251, p.279-297).
It should be noted that using a more classical definition of a species as a breeding population unable to fertively interbreed with other species, the Rockhoppers, and indeed probably all Eudyptid penguins would have to be grouped together, since successfully breeding apparent hybrids of e.g. Rockhopper and Macaroni Penguins have been reported on several occasions (White and Clausen 2002. Mar. Ornithol. 30, p.40-42).Population SizeRockhopper penguin populations appear to be declining worldwide. The Northern Rockhopper is the rarest of the Rockhopper penguins. Due to the remote locations of many colonies, census data may not be complete or accurate, yet recent data suggests that numbers are falling at some sites.Northern Rockhopper populations at Gough were estimated at 145000 in 2000 and were considered stable at the time (Cuthbert and Sommer 2004. Mar. Ornithol. 32(1), p.97-103). However, 2009 press reports citing RSPB sources suggest that numbers at Gough and Tristan are now in significant decline. A 1995 census at Amsterdam and St. Paul Islands showed that numbers on Amsterdam Island had declined compared to 1971 (58000 down to 25000 breeding pairs), whereas numbers on St. Paul had increased (4000 up to 9000 breeding pairs), possibly as a result of long-term recovery from exploitation in the 1930s (Guinard et al., 1998. Col. Waterbirds 21(2), p.222-228). |
Feeding: |
||||
The diet and foraging strategies of Northern Rockhopper penguins have been studied at Amsterdam and Gough Islands. This has revealed broadly similar foraging strategies with certain adaptations to the local environment. Diet varies not only with location but also temporally due to seasonal changes in relative and total prey availability.DietRockhopper penguin diets are highly variable and demonstrate that these penguins are opportunist feeders.The varied diet of Northern Rockhoppers at Amsterdam Island had also been recorded in an earlier study during the 94/95 breeding season (Tremblay et al., 1997. Polar Biol. 17, p.119-122). In November, 44% (by mass) squid, 31% crustaceans, and 25% fish were consumed. In contrast, in Dec. values of 15, 21 and 64%, respectively were obtained. Thus, there was a significant shift in diet from squid to fish within a relatively short space of time during the chick provisioning period. A corresponding shift from inshore to offshore foraging and towards shallower dives was noted and may have had an influence on the type of prey consumed. In contrast to the Amsterdam Island birds, Northern Rockhoppers at Gough feed predominantly (over 90% mass) of crustaceans (Klages et al., 1988. Ostrich 59, p.162-165).Foraging Ranges / Diving ParametersForaging ranges and diving parameters have also been studied at various sites during the breeding season. A study of diving behaviour of female Northern Rockhopper Penguins at Amsterdam Island provided compelling evidence for cooperative hunting behaviour in this species (Tremblay and Cherel 1999. Condor 101, p.179-185). Two females were observed diving synchronously over a period of 7 hours including 286 dives. Dives were generally initiated within seconds of each other, reached similar depths and terminated at similar times. About 92% of dives were synchronous during this period. The two birds apparently met at sea and did not leave or return to the colony together or dive in synchrony on several other days studied. It is however noted that birds often leave or return to shore in groups.
Using time/depth loggers the frequency and depth of dives can be established. At Amsterdam Island, diving of Northern Rockhoppers was studied during the guard phase in 1995 (Cherel et al., 1999. Mar. Biol. 134, p.375-385) and during the creche phase in 1994 (Tremblay et al., 1997. Polar Biol. 17, p.119-122). It is noted that figures from the two studies are not entirely comparable due to interannual variations. During the guard phase, in which only the female forages, birds were found to leave the colony at dawn, returning about 12 hours later. Transit speed to the foraging areas is approx. 7.4 km/h based on earlier studies (Brown 1987. J. Field. Ornithol. 58, p.118-125) and transit time was about 45 min either way. Most birds stayed within 6 km of the coast and performed about 550 dives. Mean dive depth was 18.4 meters, but dives up to a depth of 109 m were recorded. Average dives lasted for 57 sec, with max. of 168, and the mean surface time between dives was 21 sec. This means that birds spent nearly 70% of time at sea under water. The penguins usually descended at a rate of about 1.2 m/sec, foraged at varying speeds for an average of 28.4 sec within a narrow depth range ("bottom time"), and then ascended at 1 m/sec (3.6 km/h). The angle of descent was steeper during longer dives, thus increasing the rate of descent/ascent without changing swimming speed (which is kept at a constant, probably physiologically optimal level). The studied birds had average stomach contents of 123.6 grams (58% crustaceans by mass). As the brood phase progressed, a trend towards more numerous shallow dives was noted. Of 29 foraging trips analysed, 2 involved birds remaining at sea overnight. Diving activity was however low and shallow. Since penguins are visual predators, maximum foraging depths are attained at around noon, when the sun is at its highest position. During the creche phase, birds at Amsterdam Island were observed diving deeper and shifting from a squid- to fish-dominated diet when the early and late creche phase was compared. Mean depths of 66 m were recorded, with a max. depth of 168 m being achieved by one female. Dive depths of males and females were generally comparable.In a further study (Tremblay and Cherel 2003. Mar. Ecol. Prog. Ser. 251, p.279-297), Northern Rockhoppers at Amsterdam Island were compared to Eastern Rockhoppers at Mayes Island (Kerguelen) and Possession Isl. (Crozet) during the 1996 brood period. The birds at all sites were found to have shorter outward transit times than return transits, suggesting that they gradually move away from the colony as they forage. Overnight trips were observed at Amsterdam and Crozet, but not Kerguelen. Dive depth averages were 18.7, 33.9, 26.3 m, respectively (with 93, 104 and 85 m max. dive depths) with the deepest dives performed at the middle of the day at all sites. The average stomach contents were 107.5 g (Ams.), 150.6 g (Croz.) and 214.8 g (Kerg.) and the relative rates of chick growth varied accordingly. The amount of stomach contents corresponded to the relative levels of zooplankton biomass at the three sites.Winter DispersalAfter moulting, Rockhopper penguins usually spend extended periods at sea until returning to their colonies for the next breeding season. The location of Northern Rockhoppers during this period has not been studied. |
Reproduction: |
||||||||||||||||||||
Timing of BreedingThe breeding season commences as birds return to the colonies from their long winter foraging trips. This occurs at different times at different locations within the breeding range. Mass of the birds on arrival can be a critical determinant of subsequent breeding success (Crawford et al., 2007. Afr. J. Mar. Sci. 30(1), p.185-188; Raya Rey et al., 2007. Ibis 149, p.826-835). at different times throughout the wide breeding range of the penguin. Generally, the more northerly colonies in warmer locations start to breed earlier than those further south (approx. 10 days later for each degree C lower water temperature).Nest & Partner SelectionMale birds return to the colonies first to establish nest sites, whilst females return up to 10 days later. Interannual fidelity to nest site (>50%) and mates is considered high in Eudyptid penguins. The colonies are dense, usually in rocky areas largely stripped of vegetation. Simple nests are made using stones, twigs and other available materials and usually occupy areas near rocks or hollows beneath them. The colony is usually reached by a single or small number of tracks leading up from the coast which may be extremely steep and require the penguins formidable climbing skills.Courtship and CopulationCourtship involves advertising behaviour such as bowing and also the ecstatic display, in which all Eudyptid penguins point their heads skywards, extend their flippers and makes a series of cries with their bills wide open whilst swinging their heads from side to side. As courtship proceeds, the partners may perform the mutual display where both birds bow, quiver or perform the ecstatic display in synchrony.Copulation occurs at the nest and can be instigated by either partner. The female may squat on the nest or the male may indicate willingness by beating the female with his flipper. The male then huddles up to the female, nibbles at her neck, and pats her with one flipper. The male then proceeds to mount the female which lies with a raised head, and the male edges backwards towards the cloaca whilst beating the females flanks with his flippers. Both birds move their cloaca towards each other to make contact. The male holds its position briefly, stabilized by its flippers and often held by the neck by the female. The male then dismounts and takes a subdued hunched attitude. Both birds may then proceed to preen or simply rest for a short period.Laying / Brood Reduction MechanismNorthern Rockhopper Penguins initiate their breeding cycle about 2 months earlier than other Rockhopper Penguins breeding at lower latitudes. At Amsterdam and St. Paul, eggs are laid in early September and hatch just over a month later. Eggs also tend to hatch by mid-October at Tristan da Cunha. Within colonies, most hatching occurs within a 10-day period (Tremblay and Cherel 2003. Mar. Ecol. Prog. Ser. 251, p.279-297). |
 |
 |
|
Northern Rockhopper Penguin with Begging Chick |
Northern Rockhopper Penguin Feeding Chick |
 |
 |
|
Juvenile Northern Rockhopper Penguin |
Juvenile Northern Rockhopper Penguin Bullied by Adult |
 |
 |
|
Juvenile Northern Rockhopper Penguins with Adult |
Juvenile Northern Rockhopper Penguin with Adult |
 |
 |
 |
|
Moulting Northern Rockhopper Penguins |
 |
General Behaviour: |
||||||||||||||||||||||||||||||||||||||||
Rockhopper Penguins are considered quite fierce if disturbed and are known to peck and grab onto the legs of researchers moving through colonies. When threatened, the black crown feathers may be erected and the cheeks slightly puffed out. Rockhoppers may turn there heads from side to side and/or up and down whilst facing an adversary. This is generally accompanied by raising of flippers and sometimes cries. They may also jab towards an opponent with an open bill. If fighting ensues, birds may try to grab each other at the back of the neck and then beat each other with their flippers. Retreating birds may be followed and bitten in the hind as they flee. Aggressive behaviour is more readily displayed by males (which may account for the fact that the male is responsible for the chicks during the guard phase, where they are most vulnerable). It is noted that the Rockhopper usually only shows aggressive behaviour if it feels in danger or if its territory is challenged, especially by other males, although bullying of other birds, especially juveniles or chicks may be observed. Human beings that keep a reasonable distance from the nests and maintain calm are more likely to be met with curiosity than aggression.
The male is generally responsible for collecting nest material, whilst the female is more likely to be responsible for arranging the nest contents, especially at the egg-laying phase. Nest relief may be accompanied by loud braying (trumpeting) behaviour which may involve simple vertical trumpeting with the head pointing skywards and the flippers moving up and down, or may involve some swinging of the head from side to side, as in the ecstatic display. After this greeting behaviour the birds may switch positions at the nest (although this may not occur for some time). The penguins tend to take a relatively subdued hunched posture during this process and the returning bird gently shuffles onto the nest where it may make a couple of stepping motions whilst remaining essentially stationary before settling on the nest.Like other penguins, Rockhoppers like to congregate near sources of fresh water and can sometimes be seen bathing or drinking in streams. Cleanliness is sometimes achieved by a combination of washing / preening during short trips to the sea, during which penguins tend to stay close to the coast. Preening and allopreening also serve to maintain the plumage in good condition and remove parasites, and in the latter case additionally to reinforce the pair bond.
On land, when the penguins are not foraging or cleaning themselves, most time is spent roosting in a standing or lying position.
|
Threats: |
Rockhopper penguins have declined significantly in the last 50 years, probably largely due to competition with commercial fisheries and maybe also due to increasing fluctuations in local sea temperatures (which also impact on prey numbers and location), possibly associated with global warming. Since the populations of Northern Rockhoppers are located on small isolated islands, the species is particularly vulnerable since the birds can not relocate to other nearby sites in response to changes in local prey availability.The effect of commercial fisheries is controversial. Removing biomass from the feeding grounds of the penguins, especially near their colonies, may exacerbate food shortages in naturally occurring years with low biomass yields, leading to starvation and poor breeding success. It has however also been argued that global reductions in marine ecosystem productivity are responsible for population declines. Fisheries may certainly directly cause penguin fatalities. For example, Rockhoppers have been reported to get entangled in the nets of drift-net fisheries at Tristan da Cunha (Ryan and Cooper 1991. Oryx 25(2), p.76-79).Northern Rockhopper Penguins and their eggs have been subject to exploitation in the past. Rockhoppers have by killed for use as bait by Cray fishermen. For example, fishermen decimated numbers of penguins on St Paul Island in the 1930s. At Tristan da Cunha the practice was reported as recently as the 1970s (Richardson 1984. Cormorant 12, p.123-201). Fortunately, this appears to have stopped and Cray Fisheries appear to have little other direct impact on the penguins.A possible greater threat is global climate change. It has been hypothesized that a local drop in sea temperatures around Amsterdam Island could have led to prey migrating to more northerly waters resulting in a drop in penguin numbers. No other satisfactory explanation for the decline has been proposed. More recently, the warming of waters around Tristan and Gough is being linked to alarming declines in Northern Rockhopper numbers. Fisheries competition, predation, disease or pollution all can apparently be ruled out as responsible factors in this case. The warmer waters are considered less nutrient-rich and this may be affecting the whole food-chain in the area.Marine polution, in particular oil spillages represent a major threat to Northern Rockhopper colonies, especially since these are concentrated at only few small sites. This threat was demonstrated when the MS Oliva Bulk Carrier ran aground at Nightingale Island (Tristan da Cunha) on March 16, 2011 and spilled much of its 1500 tonnes of bunker fuel and cargo of Soya in the following days as it broke up. Oil surrounded the island and reached nearby Inaccessible Island and the main island of Tristan in the following days. A massive rescue operation was organized in the following weeks by the Islanders with massive support from SANCOB (S. African Foundation for the Conservation of Coastal Birds), which had gained experience cleaning oiled penguins following the havarage of the bulk ore carrier MV Treasure between Dassen and Robben Islands in 2000, which affected some of the largest breeding colonies of African Penguins, oiling about 20000 birds. At Tristan, nearly 4000 penguins were taken to the rehabilitation center, although nearly 2500 could not be saved. Many further birds probably were lost at sea or died before reahing the center. Preliminary studies in 2012 suggest that the breeding population is not significantly diminished although breeding success remains to be measured and long term effects of ingestion of the poisonous oil cannot yet be determined. Fortunately, the spill did not occur during the breeding season, during which the impact would presumably have been even more devastating.Predation may play a role in the sustainability of colonies. The Amsterdam Island Sub-antarctic Fur Seal population increased from 5000 to 35000 in the 1970s at a time when the penguin population was falling (Guinard et al., 1998. Col. Waterbirds 21(2), p.222-228). Seals are known to hunt penguins, although these don't usually make up a significant proportion of their prey. Sharks presumably also prey on penguins, yet little is known about at sea predation. As with many other penguin species, avian predation of eggs and young chicks is common at breeding colonies. Eggs may be taken by a variety of predators, including Skuas and other gulls. Only large predators such as Skuas take chicks. When colonies are shrinking and fragmenting due to other factors, predation may accelerate the decline, since smaller fragmented colonies have relatively more peripheral nests than large colonies and all avian predators tend to prefer hunting at the colony periphery (Jackson et al., 2005. Oikos 111, p.473-478). Interestingly, avian predation at sea was recently observed at Nightingale Island, Tristan da Cunha (Ryan et al., 2008. Ardea 96(1), p.129-134). Mature male Northern Giant Petrels were observed patrolling around the penguins landing area. The Petrels tried to grab penguins by the neck, and if successful in gaining a strong grasp would hold the penguins head under water for 5-6 minutes until it drowned. Several penguins were killed by individually hunting Petrels which either pattered across the water after penguins or lunged towards penguins that came close to them. Whilst the former technique was the most common, only 1/88 attacks observed resulted in a kill as penguins often dived to avoid the approaching Petrel. The lunging technique was more successful with 4/50 attempts resulting in a kill (although 7 birds that were initially grabbed managed to escape). One penguin was also killed after it had been dragged off the landing rock by a Petrel.Disease may also cause mortality in penguins. A number of bacterial or viral infections may affect penguin populations in general. Specific studies on Northern Rockhopper populations do however not appear to have been published |
Where To See: |
||||
Northern Rockhopper Penguins are amongst the most difficult penguins to visit in the wild due to their remote breeding locations. The most easily accessible colonies are at Tristan da Cunha. The island is about 5-6 days from Cape Town and is sporadically visited the research or merchant vessels which may offer limited passenger berths. Several antarctic cruise vessels make short stops at Tristan at the end of the antarctic summer cruise season. Landing is however sometimes prevented by harsh weather conditions.Specialized cruises have occasionally visited Amsterdam and St. Paul Islands.Captive Northern Rockhopper penguins can be seen at Vienna's highly recommendable Schönbrunn Zoo. Penguins were collected from Tristan da Cunha in 2004 in order to establish a captive breeding program. In view of the remarkable breeding successes achieved (e.g. about 10 chicks have been raised annually during 2010-12), several penguins could be transferred to Edinburgh Zoo, Scotland, in 2011. Whipsnade zoo near London, England, also has a couple of these rare penguins, as does Berlin Zoo in Germany. All images on this page were taken at Vienna Zoo unless otherwise indicated.The breeding program at Vienna Zoo involves several measures to increase breeding success. Non-paired adults and juveniles are separated from nesting penguins to reduce "jeaulousy-driven" aggression, and chicks are removed from nests after 1-2 weeks for subsequent handrearing (in 2 chick nests the larger chick is removed after a couple of days to avoid chick loss due to the brood-reduction mechanism). Zookeepers keep unnecessary disturbance to a minimum (for example not cleaning the windows between visitors and the cooled rooms housing the penguins). Light levels are also controlled to mimick those encountered by the penguins in their natural habitat.
|